
品牌 其他品牌 规格 100ul 供货周期 现货 主要用途 仅供科研
EGFR扩增和过度表达蛋白抗体VOPP1说明书别 名ARHGAP 7; ARHGAP7; Deleted in liver cancer 1; Deleted in liver cancer 1 protein; DLC 1; Dlc-1; DLC1; DLC1 Rho GTPase activating protein; FLJ21120; HP; HP protein; KIAA1723; p122 RhoGAP; RHG07_HUMAN; Rho GTPase activating protein 7; Rho GTPase-activating protein 7; Rho type GTPase activating protein 7; Rho-type GTPase-activating protein 7; StAR related lipid transfer (START) domain containing 12; StAR related lipid transfer protein 12; StAR-related lipid transfer protein 12; STARD 12; StARD12; START domain containing protein 12; START domain-containing protein 12.
研究领域肿瘤 心血管 细胞生物 免疫学 发育生物学
抗体来源Mouse
克隆类型Monoclonal
克 隆 号7H1
交叉反应 Human,
产品应用IHC-P=1:200-800 IHC-F=1:500-1000 ICC=1:100-500 IF=1:500-1000 (石蜡切片需做抗原修复)
not yet tested in other applications.
optimal dilutions/concentrations should be determined by the end user.
分 子 量14kDa
细胞定位分泌型蛋白
性 状Lyophilized or Liquid
浓 度1mg/ml
免 疫 原Recombinant human Cystatin C Protein:
亚 型IgG
纯化方法affinity purified by Protein G
储 存 液0.01M TBS(pH7.4) with 1% BSA, 0.03% Proclin300 and 50% Glycerol.
保存条件在20℃储存一年。避免重复冻融循环。冻干抗体在室温下稳定至少一个月,保持在20℃以上一年以上。当在无菌pH 7.4、抗体130M或抗体稀释液中重组时,抗体在2-4℃下稳定至少两周。
药典
EGFR扩增和过度表达蛋白抗体VOPP1说明书背景:
该基因编码一个105 kD的蛋白质,可以通过26S蛋白酶体进行共翻译处理,产生50 kD蛋白。105 kD蛋白是一个ReR蛋白特异性转录YZ剂,50 kD蛋白是NF-κB(NFKB)蛋白复合物的DNA结合亚基。NFKB是一种转录调节因子,通过细胞内、细胞外的刺激,如细胞因子、氧化自由基、紫外线照射和细菌或病毒产物激活。活化的NFKB转入细胞核并刺激涉及多种生物学功能的基因的表达。NFKB的不适当激活与许多炎症性疾病相关,而持续YZNFKB导致不适当的免疫细胞发育或延迟的细胞生长。发现了两个编码不同亚型的转录变体。[ RefSeq,SEP 2009 ]提供。
功能:
NF-κB是一种多效性转录因子,存在于几乎所有的细胞类型中,是一系列信号转导事件的终点,它是由大量的与多种炎症、免疫、分化、细胞生长、肿瘤等生物学过程相关的刺激所引发的。SIS与细胞凋亡NF-κB是由含有Rela/P65、RelB、NFKB1/P105、NFKB1/P50、ReR和NFKB2/P52的类Re-样结构域形成的同源或异源二聚体,而异源二聚体P65-P50复合物似乎是Z丰富的。二聚体结合在其靶基因的DNA中的Kappa B位点,并且个体二聚体对于不同的Kappa B位点具有不同的偏好,它们可以与可区分的亲和力和特异性结合。不同的二聚体组合分别起转录激活子或阻遏子的作用。NF-κB是由翻译后修饰和亚细胞分区的各种机制以及与其他辅因子或辅阻遏物的相互作用控制的。NF-κB复合物在与NF-κBYZ剂(I-κB)家族成员络合的非活性状态中被保持在细胞质中。在传统的激活途径中,i-KAPAB通过不同活化剂对IKKAP-B激酶(IKKs)磷酸化,随后降解,从而释放转移到细胞核的活性NF-κB复合物。NF-κB异源二聚体p65-P50和RelB-P50复合物是转录激活因子。NF-κB P50-P50同质二聚体是转录阻遏物,但与BC3相关时可以起转录激活子的作用。NFKB1似乎具有双重功能,如通过P105附着的NF-κB蛋白的细胞质保留和通过共翻译处理产生P50。蛋白酶体介导的过程保证了P50和P105的产生,并保留了它们的独立功能,尽管NFKB1/P105的处理也似乎在翻译后发生。P50结合Kappa B共识序列5’-GGRNYYCC-3’,位于参与免疫应答和急性期反应的基因的增强子区域。在与MAP3K8的复合物中,NFKB1/P105YZMAP3K8诱导的MAPK信号转导;通过NFKB1/P105的蛋白酶体依赖性降解释放活性MAP3K8。
Subunit:
NF-κB p65-P50复合物的组成。NF-κB p65-P50复合物的组成。同型二聚体;NF-κB P50-P50复合物的组成部分。NF-κB P105-P50复合物的组成部分。NF-κB P50-C-ReR复合物的组成。由NF-κB P50-P50同型二聚体和BCL3组成的复合物的组分。还与MAP3K8交互。NF-κB P50亚基与NCOA3共激活因子相互作用,通过其组蛋白乙酰转移酶活性可激活NF-κB依赖性表达。与DSIPI相互作用;这种相互作用阻止核易位和DNA结合。与SPAG9和UNC5CL相互作用。NFKB1/P105与CFLAR相互作用;相互作用YZP105加工成P50。NFKB1/P105与MAP3K8和TNIP2形成三元配合物。与GSK3B相互作用;相互作用阻止P105处理至P50。NFKB1/P50与NFKBIE相互作用。NFKB1/P50与NFKBIZ相互作用。核因子NF-κB P50亚基与NFKBID相互作用。直接与MEN1相互作用。与HIF1AN相互作用。
亚细胞定位:
细胞核。细胞质。注=核,但也发现在细胞质中的非活性形式络合YZ剂(i-KAPPA-B)。
翻译后修饰:
当翻译发生时,GRR重复后的特定展开结构促进P50的生成,使其成为蛋白酶体可接受的底物。这个过程被称为共翻译处理。被处理的形式是活性的,未处理的形式起YZ剂(IκB样)的作用,能够与NF-κB形成胞质复合物,将其捕获在细胞质中。GRR重复序列下游区域的完全折叠排除了处理。
“Ser-903”和“Ser-907”引物p105的磷酸化

Sample:
30μg(人)细胞裂解液
原发性:抗组蛋白H3(单甲基K4)(BS-28 14R)1/300稀释
次级:IrDyE800 CW山羊抗兔IgG 1/20000稀释
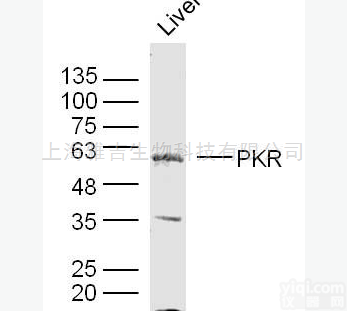
预测波段大小:17 kD
观察波段大小:17 kD
图片

多聚甲醛固定,石蜡包埋(大鼠脑);经柠檬酸钠缓冲液(pH6.0)煮沸15min后获得抗原;用3%过氧化氢阻断内源性过氧化物酶20分钟;阻断缓冲液(正常山羊血清)37℃30min;用组蛋白H3(单甲醚)孵育抗体。YL K4)多克隆抗体,未结合(BS-28 14R)于1:400在4℃过夜,然后按照SP试剂盒(兔)(SP-023)指导和DAB染色进行操作。
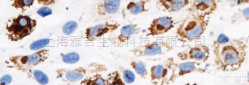 EGFR扩增和过度表达蛋白抗体VOPP1说明书
EGFR扩增和过度表达蛋白抗体VOPP1说明书
 神经母细胞瘤扩增基因蛋白抗体NBAS说明书
神经母细胞瘤扩增基因蛋白抗体NBAS说明书
 单细胞全基因组扩增试剂盒说明书
单细胞全基因组扩增试剂盒说明书
 快速DNA提取扩增试剂盒说明书
快速DNA提取扩增试剂盒说明书
 北京快速DNA提取扩增试剂盒说明书
北京快速DNA提取扩增试剂盒说明书
 WE0120型血液直接PCR扩增试剂盒说明书
WE0120型血液直接PCR扩增试剂盒说明书
 北京多重PCR扩增试剂盒说明书
北京多重PCR扩增试剂盒说明书
 单细胞RNA扩增试剂盒说明书
单细胞RNA扩增试剂盒说明书
 Anti-Tubulin-Beta抗体-微管蛋白抗体(免疫组化用抗体)Tubulin β说明书
Anti-Tubulin-Beta抗体-微管蛋白抗体(免疫组化用抗体)Tubulin β说明书
 Anti-Tubulin-beta抗体-微管蛋白抗体(免疫组化用抗体)Tubulinβ说明书
Anti-Tubulin-beta抗体-微管蛋白抗体(免疫组化用抗体)Tubulinβ说明书
 Anti-Tubulin-Beta抗体,微管蛋白抗体(免疫组化用抗体)Tubulin β说明书
Anti-Tubulin-Beta抗体,微管蛋白抗体(免疫组化用抗体)Tubulin β说明书
 Anti-Tubulin-beta抗体,微管蛋白抗体(免疫组化用抗体)Tubulinβ说明书
Anti-Tubulin-beta抗体,微管蛋白抗体(免疫组化用抗体)Tubulinβ说明书
本产品信息由(上海雅吉生物科技有限公司)为您提供,内容包括(EGFR扩增和过度表达蛋白抗体VOPP1说明书)的品牌、型号、技术参数、详细介绍等;如果您想了解更多关于(EGFR扩增和过度表达蛋白抗体VOPP1说明书)的信息,请直接联系供应商,给供应商留言。若当前页面内容侵犯到您的权益,请及时告知我们,我们将马上修改或删除。




